The Fruit World
Perhaps the closest grouping to nuts is that of fruits. Although most people are familiar with only ten or twenty different fruits, there are at least 10,000 different plant species which can be regarded as fruit plants. Like nuts, they extend across the whole range of plant species, and also extend over the whole planet. Some plant families include both fruits and nuts.The fruit families give exactly the same distribution picture as the nuts. Figure 6.1 shows the distribution of the Sapotaceae, of which perhaps the best-known representative is the Sapodilla or Chico, Manilkara zapota. As well as producing a delicious fruit, this species is also the source of chicle, the base for chewing gum. The family produces many other fruits and plant products, including some where the masses of sweet edible flowers are the crop (Madhuca), and others with seeds high in edible oils (Butyrospermum, Shea Butter).
This distribution shows all the features which we have come to expect from an Earth-expanded family. The Sapotaceae originated in Gondwanaland, and the basis of the distribution is a band extending round the Earth through central America, central Africa, across the Indian Ocean and India through the Indonesian islands and northern Australia, then back across the Pacific to America.

It seems reasonable that the distribution was once a fairly uniform band running right around the Gondwanan tropics. This uniformity has been disturbed by the "centrifugal" movement of domains, carrying species as far north as Florida, the Bahamas, Morocco, and Taiwan, and as far south as Chile, New Zealand, and South Africa. The typical mechanisms of microdomain shuffling and flight are clearly active.
The occurrence of the same genera on either side of the Pacific (eg Chrysophyllum, the Star Apple genus), and even stretching right across from Central America though Asia/Australia over to southern Africa (Manilkara itself) show how recent and rapid the opening up of the Indian Ocean and the Pacific Ocean were.
I have already mentioned that the puzzle of the coconut (whether it originated in southeast Asia or on the northwest of South America) can be explained by assuming the original species distribution was split apart by Earth expansion. A similar case occurs with a fruit from the persimmon or ebony family, Diospyros ebenaster, called Zapote Prieto.
This family, the Ebenaceae, has a very similar distribution to the Sapotaceae, with a particularly strong distribution band over from Central America through Asia/Australia to central and southern Africa. The Diospyros genus itself (it also includes the ebonies) runs right across, and is notably strong, both in the Philippines (38 species) and in Mexico (11 species).
One species, D. ebenaster, occurs in both areas [Standley, 1924], and there are good separate cases for assuming that it is native in Mexico and it is native in the Philippines. The obvious conclusion is that it is native to both. Even if this particular conclusion is wrong, say because two different species of Diospyros happened to evolve to be almost indistinguishable in the two areas, this does not explain how the strong concentrations of Diospyros species came about, so far apart. Or how there is a further concentration (23 species) in southern Africa.
Since starting the work on which this is based, I have examined the distributions of something approaching 20,000 different species of fruit plants and their relatives, keeping a particular eye out for anomalies. I have been unable to find a single example of a plant which glaringly contradicts the principles outlined.
Such mild counter-evidence as exists is usually explicable as due to either to long-distance bird migration, as with some small berry species (eg Vaccinium, the blueberry family), or to really ancient lineage in the case of a few genuinely world-wide families such as the Fagaceae (oaks, beeches, chestnuts) and Salicaceae (willows), which have representatives in both northern and southern temperate areas. The usual explanation of a plant with a strange distribution, that it "was introduced by man", in fact seems to have been invoked quite unnecessarily and wrongly in many cases.
Proposition 6A
Many cases of plants assumed to introduced to have been
introduced by man, to explain their occurrence, are as readily
naturally explicable through expanding-Earth principles
The Families of Plants
When the distribution of almost any plant family is examined, a good case can usually be made for assigning it to either Laurasian or Gondwanan origin. Table 6 lists some of the world's major plant families under one of these two headings. The listed families include the majority of the Earth's tree species.Table 6. Gondwanan and Laurasian Plant Families
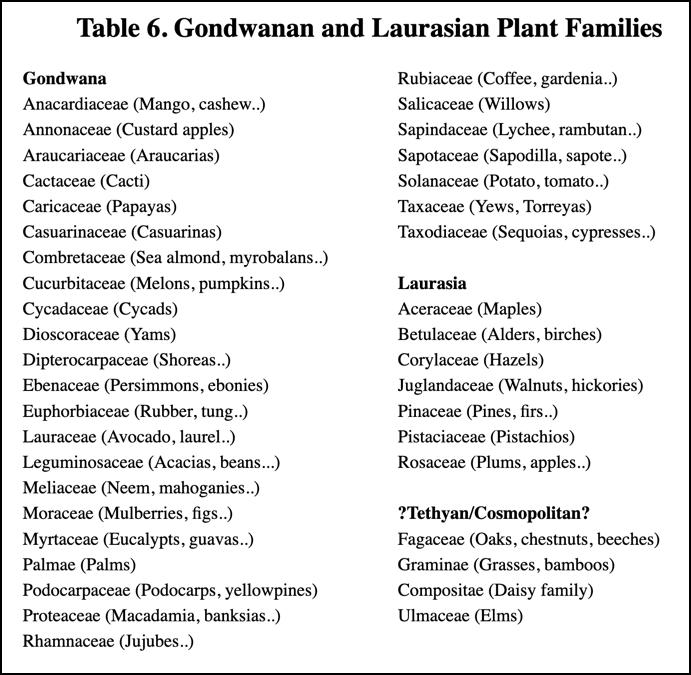
Some of these families contain hundreds, or even thousands, of species. With these numbers they may be very widespread, and in a typical enumeration (such as in [83]) they may be described as "world-wide" or "pan-tropical".
Even so, when one of these families is examined in detail in the light of the domain movements which have been identified, it usually falls in one camp or the other. Most of the genera which look to be exceptions to the rule for their family can be explained either by these domain movements, or on the grounds that the family is an old one which was actually split apart when the Tethyan girdle was formed.
Then again, the exceptions may be purely a matter of classification. The pistachios, clearly a Laurasian genus, were usually classed in the Gondwanan mango and cashew family, the Anacardiaceae. More recently they have been separated off into their own family, the Pistaceaceae, which moves them out from being an exception. Currently the Fagaceae includes the Laurasian oaks, chestnuts, and beeches (Fagus), but also the Gondwanan southern beeches (Nothofagus) and tropical chestnuts (Castanopsis). Perhaps closer study will separate off these latter genera into a separate family.
We have widened the scope of our study from nuts, to all fruits, then to all plants. Now we can widen our view even more, to encompass the other great Kingdom, the animals.
Lower Animals
Some of the earliest animals which are believed to have lived partly on land, the amphibians, are assumed to have evolved from early fishes. The link between these two classes is the group of lung-fishes, fishes in which the swim-bladder has altered to perform some of the functions of an air-breathing lung.Living lung-fishes are today found in three continents -- Australia, Africa, and South America, together making up the bulk of Gondwanaland. Lungfishes live at the boundaries of land and fresh water, usually in the banks of rivers which may become seasonally dry. We will return to the matter of the land/water boundary in NU010.
Further up the evolutionary scale, when we look at the crocodiles and alligators, we can see that these are exclusively Gondwanan. Africa, Australia, South America, Southeast Asia, and Florida -- that is where the crocodilians are, in almost every part of the former megadomain. Even turtles, marine amphibia with the obvious opportunity to spread their range, show a similar pattern.
Marsupials
As is well known, the vast majority of marsupals living today are native to Australia. A few are found in the islands to the north, such as New Guinea. New Zealand has none.The major exception to the Australian-only distribution is the opossoms of the Americas. There are only two American genera, one of which contains only one species -- the Yapock. This sole strange aquatic and web-footed representative of the genus Chironectes exists from Guatemala down to southern Brazil.
Most of the species in the other genus, Didelphys, are also concentrated in tropical America, that is, northern South America and southern Central America. However, by the chances of European settlement, the best-known one is the Virginian Opossum, Didelphys virginiana. Like its Australian possum relatives, this animal has adapted well to modern civilization and can be found happily living in the roofs of houses.
Now unlike plants and lungfishes, these marsupials are clearly mobile creatures, and the fact that opossums are common over much of the United States is a reflection of that. Nevertheless, I will show that this mobility is much more limited than is generally assumed.
In the very first Proposition in this suite of articles, number 2A, I suggested that the actual rates of spread of plants are much less than their potential rates. A similar proposition can be formulated for animals too.
Proposition 6B
Actual rates of spread of animals are usually much less than
the potential rates of spread implied by the mobilities
of individual animals
As for the mechanism behind this, this has already been given in Proposition 2B. Like plants, animal species do not expand their range because they are unable to overcome ecological pressures.
Some of the conclusions to be drawn from the above are obvious. Marsupials exist in greatest quantities in widely-separated Australia and northern South America because these two continents were once in contact. The same is true of New Guinea and Central America -- again, look how excellently New Guinea fits against the Central American coast, reproducing every in with an out, and vice versa.
The usual reason given for the virtual restriction of marsupials to Australia is that they have survived there in isolation because they lacked competion from more highly-evolved (and implicitly "more efficient") creatures in the rest of the world. On examination, this view can be seen to lack credibility. The marsupials of the Americas have shown adaptability and resilience in maintaining and even extending their range in the face of ecological pressure from "more evolved" creatures. The marsupials aren't elsewhere because they never were.
Proposition 6C
Marsupials evolved in the Australian and South American
domains when these were in contact, and were not cut back to
these areas because of competition from "more evolved"
creatures
In opposition to this could be quoted the matter of fossils. Remains believed to be those of fossil marsupials have been found in England and in France. This is not a serious objection -- see Proposition 5G. The areas containing these fossils in England and France are almost certainly microdomains which have been separated from their fellows on the other side of the Atlantic.
Why should marsupials be found in North America at all? The answer is to be found in those north-thrusting domain belts moving up on both sides of the North American continent, domains which were once part of Gondwanaland.
It is no coincidence that the common U.S. opossum, Didelphys virginiana, has Virginia in both its common and scientific names -- Virginia was once part of Gondwanaland. Neither is it a concidence that the scientific name of the American Persimmon, the country's sole representative of the Gondwanan family Ebenaceae, is Diospyros virginiana.
Other Mammals
When other mammals are looked at, it is a question of more of the same. Most groups of mammals can readily be classed as of either Gondwanan or Laurasian origin.Consider bears. These are still virtually confined to Laurasian domains, the small extension down into parts of southeast Asia is a credible natural spread. The same is true of our oldest domestic pet, the dog, and its relatives the wolves.
Our other common house pet, the cat, is Gondwanan. Cats were pets in ancient Egypt, one of the closest bits of "Gondwanaland" to ancient Greece. Tigers in India and Indonesia, lions there and in Africa, pumas in South America, are all relatives calling Gondwanaland home. The north American cougar presumably rode up on the western microdomain shuffle belt.
Other Gondwanan families include the elephant -- now we have a natural explanation for these animals in both Africa and southern Asia. Probably the pig, and certainly the rhinoceros (with representatives in Africa, India, and Indonesia), come in here. The positions of the deer, sheep and camel families need closer analysis to make their positions clear.
Primates
The primates are clearly of Gondwanan origin. The most primitive primates are the lemurs, lowly monkeys confined to the island of Madagascar. Their more evolved relatives extend in the familiar band up through Africa and across to India and southeast Asia.The lower apes, such as the monkeys, exist also in the tropics of South America. They differ from the others in having prehensile tails. Although these are obviously Gondwanan, it is not clear whether they are connected with the monkeys of western Africa or with those of eastern Malesia. I would suspect the former, but the sort of detailed genetic analysis which is now becoming possible [72] will probably settle the question.
The higher apes conform to the pattern, with chimpanzees and gorillas in Africa and gibbons and orang-utans in India and southeast Asia. As for the "naked ape", man himself, the position is made more complex by his ability to use intelligence to overcome ecological pressure and spread very widely (or perhaps it demonstrates the rule, with no competing intelligence to stem the spread). However, man is widely assumed to have originated in Africa, which fits in. Of this, more later.
But when it comes down to the races of man, the same pattern repeats itself, now rather uncomfortably. The original inhabitants of Madagascar are known to be closely related to the Malays, even having languages in the same group. Of course, the Malays are fine seafarers, and the link can be explained through this. But what about the Negrito people of the southern Philippines, who have been said to be genetically identical to the African Pygmies?
Even I am not bold enough to make a Proposition out of this one -- the times are apparently all wrong. Man himself is supposed to be no more than 1-2 my old, with the earliest man-like apes going back no further than 5 my, and so far no-one has suggested that the Indian Ocean opened up in as short a time as this.
The Big Birds
These give the usual pattern. The large flightless birds, the ratites, are very Gondwanan. The ostrich in Africa, the emu in Australia, and the rhea in South America are not only similar and related, but even fill similar ecological niches -- they are grazing animals. In northern Australia and New Guinea the cassowary is a relative, as is the kiwi in New Zealand.It is interesting that the ostrich once extended right across north Africa, and into Arabia, even in recent historical times -- perhaps there are still a few left there. Arabia is an interesting domain to locate.
What is also interesting is that these ratites had giant relatives, now unfortunately extinct through the hands of man. The huge moa, standing 3.6 m high, is believed to have been wiped out by the first human settlers in New Zealand, and the same fate may have overtaken the gigantic Aepyornis of Madagascar. At least we need not blame ourselves for the loss of probably the most massive bird of all time, the Dromornis of central Australia, which weighed 500 kg; this apparently died out over 10 my ago.
Some modern birds routinely undertake annual migrations of up to 10,000 km, and it is perhaps understandable to assume that they must be exceptions to the restraints on spread which apply to normal creatures. Even so, the extent of this exception is not as great as it could be.
We have already seen, in NU002, how the finches of the Galapagos evolved into a number of different species, even though they were normal flying birds, and were not distantly separated. A study of the basic genetic material (DNA) of birds of Australia and elsewhere [72] has shown that the birds of Australia are related much more closely to each other than they are to apparently similar birds from elsewhere. This is in contrast to previous assumptions, based on similarity of form, so that the magpies of Australia are closer to Australian parrots than they are to European magpies.
Marine Life
With all the oceans of the planet linked, we might expect the easy transport afforded by floating about in ocean currents to have made ocean life fairly homogenous everywhere. Such is not the case.One of the reasons for this is that a high proportion of marine life is restricted to shallow water, which lies mostly at the sea/land boundaries. As usual, this is because of ecological pressure -- away from the seashore, in deeper water, conditions change rapidly.
This means that the boundaries within which sea plants and animals are evolving, what we have referred to as isocons, are usually quite thin strips running parallel to seashores, instead of huge ocean areas. The actual "surface" areas of these isocon envelopes are often quite small in comparison to those their land analogues. And, of course, the only places where spreading can easily occur are at the two ends of the strips, rather than on all sides.
Proposition 6D
The majority of marine creatures are ecologically restricted
to shallow off-shore waters, and so inhabit long thin ecological
strips of relatively small area
Of course many marine creatures have free-floating stages, and so have the capacity to spread widely as far as their mobility is concerned. Even so, the same pattern of species is found as on land; creatures such as crabs are around almost all the world's seashores, but the species are different in each area.
Figure 6.2 (from [67]) shows the distribution of eelgrass, Zostera marina. Eelgrass is one of only 49 species of plant, collectively called seagrasses, which are true flowering plants which have adapted to live in the sea. They produce flowers and pollen and seeds, in the normal plant cycle, all entirely submerged under the water.
 Fig. 6.2. Distribution of eelgrass, Zostera marina
Fig. 6.2. Distribution of eelgrass, Zostera marina
Most of the seagrasses are found in the southern hemisphere, particularly around Australia. Eelgrass is one of the few northern species, although it has relatives in the south, and thus has a common ancestry with them (Proposition 3A). The heavy black lines along the Atlantic parts of the distribution are where the plant was affected by a devastating disease in the 1930's.
The points to be made from this map are that eelgrass has a clear Laurasian distribution, and that even though it is a marine plant, its current distribution was presumably brought about by Earth expansion after the species had evolved and separated off from a common southern-hemisphere stock.
Notice that the two parts of the distribution are no longer connected, although they must have been at some time in the past. Also that the vector carrying the wasting disease was able to cross the Atlantic (and hence had a different ecopressure boundary to eelgrass itself), but it did not reach into the Mediterranean (or across to the Pacific).
We have seen that the consequences of Earth expansion have been effective across the whole spectrum of animal and plant life, not only life on land, but life in the sea as well. Now we have enough evidence to go on and see how far we can reconstruct the Earth as it was in former days.

(Full list of references at NURefs)
[67]. Erik Rasmussen, Erik. The wasting disease of eelgrass (Zostera marina). In: Seagrass ecosystems, a scientific perspective. Marcel Dekker, 1977.
[72]. Charles Sibley. Reconstructing bird phylogeny by comparing DNAs. Scient American/ Feb p86, 1986.
[83]. J C Willis. A dictionary of the flowering plants and ferns. 8th ed. Cambridge University Press, 1973.
NU007: Putting The Earth Back Together
NU005: How The Earth Fell Apart
Go to the NUSite Home Page

Version 1.0, in printed edition of "Nuteeriat: Nut Trees, the Expanding Earth, Rottnest Island, and All That...", 1989.
Version 2.0, 2004, PDFs etc on World Wide Web.
Version 3.0, 2014 Sep 10, Reworked from "Nuteeriat" as one article in a suite on the World Wide Web. V. 3.1, Adjusted, 2025 May 31.
V. 4.0, Adjusted for *, 2025 Sep 30.