It would be nice to present here the complete picture, but, alas, that is quite impossible. It will require years of painstaking work by many people, plus probably the need to go out into the field and accumulate new evidence, before all the arguments will be settled and we feel that we really have got the thing right. All we can do here is make a start on the new area of investigation called Domainography, by putting together some working rules and giving a few possible answers.
The Rules of Domainography
The first aim of the approach must be to keep it as simple as possible -- we will expand on this further in NU017. When the broad outlines are established, then will be the time to find the exceptions to the rules and possibly to refine or extend those rules. At the same time, we know that the general rules will be extracted from a mass of individual, detailed examples. What general rules should we have? We can start off with the following:RULES FOR HOLODOMAIN RECONSTRUCTION
Rule 1. Larger landmasses are composites of many domains.
Rule 2. As domains broke up, the relative positions of the pieces stayed the same, while their distances apart increased.
Rule 3. There were no large rotations of domains.
Rule 4. Smaller domains have moved further than larger ones.
Rule 5. Domain splitting was according to the normal behaviour of materials.
Rule 1 is a basic assumption. Rule 2 simply means that if Domain 26A was to the east of Domain 26B before Domain 26 split, then 26A will always be more or less to the east of 26B, even though they may end up hundreds or thousands of kilometres apart.
Rule 3 is an interesting one. At this stage it is purely a rule-of-thumb. There is, in fact, a physical law, called the Coriolis Force, which does cause rotation of bodies moving away from or towards the Equator (this determines the direction of rotation of cyclones and the way water swirls down the plughole). But all instances of domain rotation observed to date seem small enough to make their existence questionable.
We might expect this rule to be broken when a smaller, irregularly-shaped domain impacts against a larger one at an oblique angle. India looks as if it might have rotated a few degrees anticlockwise when it ran into Laurasia and made the Himalayas. However this apparent effect could be illusory, caused by differences in the angle of viewing a landmass at different latitudes.
Rule 4 is also an observed rule of thumb, derived from the distributions of plants and animals which we have already examined. For Rule 4 to apply, it obviously must be physically possible; there is no way a microdomain is going to force its way through a megadomain it has run into, to keep up with its fellow which was off to one side of the megadomain.
Rule 5 is an important one, with many consequences. It just says that all domains are made up of masses of rocks which have known physical properties, and the domains will obey normal physical laws of every sort. The fact that we may not have a complete picture of some of these laws does not affect the point.
We already invoked Rule 5 when it was pointed out that the deep ocean trenches could not have been formed by pushing one slab of rock against another (see Proposition 3C) -- such an act could not create a gap between them. Another consequence is that the new boundary formed when a domain was torn apart could not be especially complex.
Such a new boundary might well vary from very clean and straight (such as that of the coast of Israel and Lebanon) to somewhat irregular and jagged. But normal physical forces would not permit a break to be formed in the shape of a deeply concave bay, such the Gulf of Carpentaria on Australia's north coast. This must be a composite of some sort, there must be at least two domains involved in its formation. This also applies to any other 'complex' coastline or domain boundary, complex on a scale beyond that of erosional effects. We have already seen, for example, how major peninsulas involve at least two different domains. This is important enough to extract as a new rule:
In addition, there is one more rule which has justification to be explained later (NU008):
The last item in our armoury, before we look at actual re-assembly, is a useful technique.
The Rubber Band Technique
There is a convenient visual approach we can use in this. Look again at some of the plant distribution maps given in Chapter 4. Assume all the land areas have been divided up into domains.Now regard the distribution boundary as a rubber band, and shrink this band down. It will tighten, and pull together all the domains which were once connected by the early distribution range of the plant genus or species. This will give a preliminary picture of how the domains involved once fitted together on the pre-expanded Earth.
This technique can be used not once for each domain, but hundreds of times -- once for each plant species, genus, or family represented on it. It can even be used for 'races' within a species, and if the genetic patterns of individuals are available, say through the isozyme analysis method, it can be used with them too.
Of course this technique will also drag in a ring of domains which do not form part of the original distribution, but instead are just current neighbours of original domains, neighbours onto which the plants have spread through natural dispersion processes. These neighbour domains need to be discarded.
The width of the neighbour ring to be discarded depends on how specific the level of plant description is, and what the likely rate of natural spread is judged at. To deal with the last part first, we will set up the last of our rules, which is an attempt to put numbers to Proposition 2A:
At first sight, this estimate seems incredibly low. One metre per year is only 1 km in a thousand years, which seems very little, however it is also 1000 km in a million years, and that is quite a lot. Many plant species lie entirely within a range which is less than 1000 km across. We have already suggested (Proposition 2I) that the half-life of a species is around one million years, so, in fact, this figure does seem to be of at least the right order of magnitude.
On the matter of plant description specificity, the broadest grouping, that of plant family, is too broad to be of much help here. Most modern plant families, such as the Fagaceae (which includes the oaks, chestnuts, and the northern and southern beeches) date back to the early part of the Cenozoic Era, 50-70 my ago. This would give them the opportunity to spread 50-70,000 km, much more than the distance right round the Earth (40,000 km). However, again it does seem of the right order of magnitude, if anything favouring a lower figure than that in Rule 8.
We have never put a figure to the half-life of a plant genus, such as Araucaria, but it would be somewhere between the species half-life and the known age of the family it is part of. Suppose we put a value of 10 my to this half-life. Then the average distance we would expect a genus to spread naturally would be 10,000 km, and the most a species would extend would be 1,000 km.
Although the Rubber Band technique presented above was a simple visual technique, it could in fact be developed into a computer approach, in which the width of the discardable neighbour rings could be easily varied to see the effect on the final re-assembly. I suspect that in many cases the width of these rings could be cut down to less than the distances implied by the above estimates without affecting the validity of the technique.
For example, look again at Figure 2.1, which shows the distributions of seven different Canarium species (the map for the whole genus is Fig. 4.7). It would be my contention that all the areas within the range of the most widely-spread species, number 1, C. littorale, were once joined and the species has ended up in the different major islands and peninsulas principally through Earth expansion. This is even though the name of the species means it is a sea-shore plant, and the seeds are designed to float and could well cross from island to island, making this is a worst-case example.
The current range of the species is about 2000 km across. If the islands were all pushed together by tightening the rubber band, it would drop down to a width of around 1000 km -- which is equivalent to an overland spread from a single centre of only 500 km.
Now we can look at different areas of the current world, suggesting how they may have been related in past times before the Earth expanded, and applying all the above rules. As usual, the main evidence comes from the distribution of plant species, genera, and families.
Australia
Australia appears to contain at least two megadomains, probably with a common boundary running roughly north-south through Lake Eyre and down to Spencer Gulf. In addition, it has either a long thin wedge or a microdomain shuffle belt running down the east coast, to the east of the Great Dividing Range.The east coast geology looks more like a shuffle belt. Certainly there is a marked movement of species down the east side of Australia. Typical 'tropical' genera which extend as far south as northern New South Wales include Canarium (Fig. 4.7) and Carissa. The Bunya Pine (Araucaria, Fig. 4.4) of southern Queensland (but with an outlier in North Queensland, and most of the family in New Guinea) has also presumably been carried down on this belt.
The whole of the northeast coast of Queensland has a complex rain-forest flora with strong links with the northwest coast of South America. Virtually every major family present in that area of South America has close relatives in northeast Queensland, often in the same genus (Pouteria, Syzygium, Endiandra, Litsea, Cryptocarya). The excellent fit of eastern Australia against western South America has already been remarked upon, the evidence is overwhelming that these two areas were once in contact.
In fact the evidence for this contact is much stronger than that for the Africa-South America contact which has been taken for granted since the earliest days of the Continental Drift idea, with its origins with Francis Bacon, right back when the Spanish and Portuguese explorers first mapped the southern Atlantic coasts.
This contact is a major distinction between the Continental Drift approach and its later Expanding Earth derivative. In the former, Australia is invariably placed at the opposite end of Gondwanaland to South America, and usually has adjacent New Guinea poking out into an unmapped sea. In the latter, Australia and New Guinea nestle snugly against South America and Central America, and the strong family links are easily explained.
Although the Myrtaceae family which contains the Eucalypts is well represented in South America (Eugenia has representatives in both domains), the typically Australian Eucalyptus is not. In fact the eucalypts are not especially common in the northeast Queensland rainforest, and this domain may have been separate from the rest of Australia at one time. Certainly the eucalypts appear to have evolved after the two megadomains separated. This separation presumably took place before the migration of monkeys from west Africa into northern South America.
The whole of the northern coast of Australia shows strong floristic links with adjoining areas of Asia. Nut-bearing genera such as Canarium and Terminalia are typical examples, although these extend well beyond the immediate area. One interesting genus is Horsfieldia, a relative of the nutmeg. This has around 80 species scattered through southeast Asia, including Indonesia and southern China, and one species, H. australasica, found only in the Northern Territory and the Gulf of Carpentaria. It has been recommended for development as a commercial nut species [39].
Australia also has species of Myristica (nutmeg) and Zizyphus (jujube or chinese date) matching those found in southern Asia as far north as the Himalayas. The jujubes are notable for the fact that they thrive in arid inland areas and could not be expected to cross sea barriers.
The west coast of Australia shows typical features of microdomain flight, as we have seen with Rottnest Island and various peninsula formations. As far as more distant links are concerned, a major feature is the strong connection between southwest Australia and South Africa, as shown especially through the Proteaceae family. These, typified by the beautiful Proteas of South Africa and the Banksias of southern Australia, provide probably the strongest southern link between the two continents; there are hundreds of species in each. On the nut side, the Van Riebeck almond, Brabejum stellatifolium, of South Africa, is closely related to the woody pears (Xylomelum) of Western Australia.
There are plenty of links between northwest Australia and eastern Africa, as for example Canarium (Fig. 4.7). Recently a new species of blind hunting spider was discovered in the Cape Range peninsula of the west coast [38], and its nearest relative was noted as originating in southern Africa.
On balance, it seems likely that southwest Australia was once in actual contact with what is now southeast Africa. The fit is quite good. It is possible that Madagascar intervened, but this has probably come down from further north -- see later. The separation of Australia and Africa must have been of middle age -- not ancient, but before the evolution of the eucalypts.
It should be pointed out here that if Australia was in contact with Africa and South America on the west and east, and these two other continents were themselves in contact along their Atlantic coasts, this means a very tight fit of these three continents around the South Pole must have occurred, if their shapes were the same as now. This would have left very little room for Antarctica, even the small one which we have seen is actually there (Proposition 5B). All this is evidence both of re-seaming and domain shifting in the megadomains. It also implies a pre-expanded Earth somewhat smaller than that usually assumed, closer to 50% of the present diameter instead of the 55-60% sometimes assumed.
Direct evidence of modern plant links between Australia and Antarctica is virtually lacking, because Antarctica proper supports no higher plants, and there are no major islands between Australia and Antarctica. As for the island State of Tasmania, this is a typical island microdomain, with a flora easily represented as having evolved from a much earlier 'tropical' assembly, carried well to the south. It has Nothofagus, the Southern Beech, which it shares with the New Guinea highlands and with New Zealand and southern Chile. It has one representative of an ancient conifer genus, Dacrydium, most species of which are found in Malaya and Indonesia (Malesia), but which has representatives in New Zealand, New Caledonia, Fiji, and Chile.
More Propositions
The distribution of Dacrydium is a good starting point to generalize some inferences into formal propositions. No conifer is noted for its ability to spread by sea, and it appears impossible to explain this distribution rationally except by assuming that all the above areas, now tens of thousands of kilometres distant, were once linked. The distributions, and the current characteristics of the species involved, follow naturally from the following Propositions:Proposition 7A
In the early part of the current (Cenozoic) era, 50-70 my ago,
the immediate ancestors of most of our current plant genera were
evolving out
Proposition 7B
This evolution took place in one or more 'equatorial bands'
of physically interlinked domains extending right round the
Earth, with easy spread of species along the bands
Proposition 7C
The climatic conditions in these equatorial bands were
closer to those of currently temperate areas than to modern
tropical ones
One note to this last Proposition -- it is based on the twin observations that there are a few families with representatives of the same (or closely-allied) genera in both southern-hemisphere and northern-hemisphere temperate areas, and that when representatives of these genera still exist in equatorial areas, they appear to have mostly retreated to the cooler uplands.
Examples include the Beeches, represented in the north by Fagus and in the south by Nothofagus, with the latter having species in the uplands tropics, the willows (Salix), and the Junipers (Juniperus). Most instances of these dual-temperate genera fall easily into this pattern. The inference is that the families involved are the more ancient ones, and the corollary is that typical 'tropical' families are of more recent origin.
Proposition 7D
Typical 'tropical' plant families are of relatively recent
origin, less than about 50 my old
New Zealand and the Pacific Islands
In spite of its clearly temperate location, New Zealand is noted for the 'tropical' appearance of its plants. We have seen that it has one of the most southern representatives of the Sapotaceae, it also has representatives of Eugenia, Aleurites, Freycinetia, and a palm, Rhopalostylis, as well as a host of other 'tropical' genera. However, it has no native eucalypts or wattles (Acacia, and evidently was not attached to Australia in recent times. It has very few deciduous trees, another indication of non-temperate origin.In fact, in spite of the modern separations, New Zealand has links perhaps closer to Chile than to Australia, particularly in the South Island -- it is quite possible that the two major islands were not so closely linked in the past. It seems likely that both islands have moved south relative to larger landmasses, possibly to different extents.
The islands of the Pacific show all the features of equatorial-band development and domain flight which we have come to expect. These islands are not tricky to reassemble into their pre-expansion conformations with a high degree of precision, purely by repeated rubber-banding of all their native species. The general picture is unmistakably one of a tropical development band stretching from Indonesia across to Central America, split into thousands of smaller parts, with a few islands on the northern and southern limits of the band showing equatorial flight into more temperate areas.
In the rubber-banding, it is important to question (Proposition 6A) the usual assumption, that many species have been introduced. The Hawaian group, in particular, has many species assumed to have been introduced, but which fit excellently into the expansion pattern. An example is the legume Leucaena, with 50 species in tropical America and two others stretching across through Hawaii to Polynesia. In fact is is probably more sensible to reverse the procedure, and determine whether or not a species has been introduced by seeing whether or not it 'rubber-bands' in a normal way. This test is quite independent of any historical evidence, such as the assumed spread of the coconut across the Pacific through the canoes of the islanders.
Proposition 7E
The match of a plant's current distribution pattern with other
local isocons gives evidence of whether or not the plant was
introduced by man
South and Central America
South America appears to contain two large domains, the bigger mostly made up of Brazil south of the Amazon, and the smaller the Guiana Highlands area to its north. It also has the most prominent domain shuffle belt in the world running down its west coast. There is an area of accreted islands or re-seamed domains east of Panama and extending over much of Columbia and Venezuela.We have already seen how the western shuffle belt has acted as a raft to carry species southwards. A good example is the Pygmy Coconut, Jubaea spectabilis, which is confined to an area of central Chile (Fig. 4.14). Of course palms are typically tropical, but this one is the most southern and cold-resistant palm of all, able to withstand snow.
Like all palms, Jubaea is of Gondwanan origin. There are, however, a number of species of Laurasian origin which have managed to jump onto the shuffle belt. Figure 7.1 shows the distribution of modern and fossil occurrences of the walnuts, Juglans. It is taken from Berry's classic book [9].
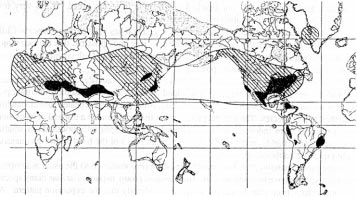
Notice that the modern walnuts occur right across what we can regard as the South Laurasian Equatorial Band -- they are obviously Laurasian -- and that the fossil occurrences (clearly somewhat sweepingly brushed in) extend up to Greenland. We know now why this is so. Notice also that occurrences of modern walnuts are shown in the Caribbean and extending down the Andes as far as Ecuador and Peru. These are also readily understandable, typical microdomain band shuffling.
But look again, and notice a large area shown for walnut in eastern Brazil, on the coast. This does not fit our pattern at all. I believe that it is an error.
Everyone can make mistakes -- there may well be some in the maps I have put together myself -- but the important point here is that we can use the reasoning developed so far to predict that this walnut placing is an error, and that prediction can be checked. Ability to produce predictions, and their verification, is one of the most important facets of any scientific theory.
Another interesting point which emerges, if you look at the various plant species of northern South America, is that species and genera on the Amazon Basin side are often quite different to those on the western coast. As an example, look at Figure 7.2, taken from Mitchell [52], which shows the distribution of species of Anacardium, the cashew genus.
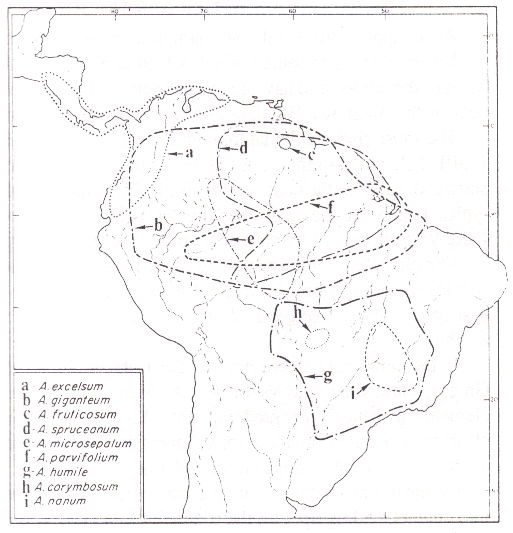 Fig. 7.2. Distributions of some species of Anacardium
Fig. 7.2. Distributions of some species of Anacardium
The Anacardiaceae family which includes the cashews has representatives across the tropics, including the Australian cashew, which however is no longer placed in the genus, but in Semecarpus. The former is now restricted to 10 species, the nine shown in Fig. 7.2, and the cashew nut itself (A. occidentale), which occurs in the same area as the rest. However, the closest relatives to Anacardium are not species of Semecarpus, but the three species of Fegimanra which grow in tropical West Africa -- again fitting the pattern precisely.
The point being made here is that neither the cashew genus itself, nor its close relatives, occur on the Andes coast of South America except in the extreme north, so it is presumably a fairly recent passenger on the shuffle belt. The genus does extend much further southwards on the east of the Andes, right down to northern Uruguay.
The walnut and cashew maps also help to define the status of the Caribbean islands and 'mainland' Central America. The walnut does not occur on the mainland, only on the islands. The converse is true for the cashew. Clearly there has been mixing of Laurasian and Gondwanan species in this zone, but the general position is consistent with the assumption that both parts are made up of a collage of microdomains which formerly lay right across the boundary of the two megadomains -- part of what we have called the Tethyan Girdle. Detailed reconstruction involves only repeated application of the rubber-banding technique.
It also seems that the present land bridge between North and South America is not particularly old, and previous land connections may have been further east. Mainland Central America is a clear aggregation of different domains, showing very obvious peninsulas and large lakes formed through the chance conjunction of different island outlines. The Galapagos islands off the coast of Ecuador have cacti (Opuntia) closely related to those of Mexico, and have been shifted southwards.
The relation of South America to Australia has already been discussed. The link between South America and Africa is obvious and inarguable. But even here, we can perhaps add a little, on relative ages.
The Brazil-West African connection is strong. We have seen it with the oil palm (Fig. 4.3) and with the cashew (Fig. 7.2). The connection can be shown again and again, through different plant families; a further good example is the custard apple genus, Annona. This is most widely represented in northern South America, although there are a few species right across tropical Africa and down the east coast. There is also a close relative, Asimina, which extends from Florida up the east coast of North America -- of this more later.
But when you look further south, for connections between Argentina, Uruguay, and southern Africa, the links are quite weak. This can be seen from the Proteaceae map, Fig. 4.1, which has a notable blank area in southeast South America. The inference is that the southern gulf between Africa and South America opened up well before the northern separation.
Africa and the Indian Ocean
Africa appears to be a relatively simple construction. It contains a number of large domains -- these could be represented as Nubia, Congo, Kalahari, Somali, and North and South Sahara. Most of these appear to have undergone some shifting and re-seaming, giving rise to features such as the Adamawa Highlands between Nigeria and Cameroon.The most obvious domainographic feature of Africa is the east coast. This is another shuffle belt, not as dramatic as that of South America, but considerably more complex in the sizes and nature of the domains involved. These have all moved southwards, leading to such typical features as the many long, thin lakes. The Ethiopian Highlands and those surrounding Lake Victoria could probably be accorded the status of separate domains.
The plant distribution pattern is fairly plain -- most families have bands of genera and species extending along the tropical lines of latitude, with extensions southwards along the east coast. The coffee genus, Coffea, is a typical example, as are the African examples of Annona.
In the north and east we run into the Tethyan Girdle, in this case including Spain, Italy, Greece, Turkey, and the Mediterranean islands, all mixtures of Laurasian and Gondwanan elements, with the former predominating. From the plant distributions, the Canary Islands are clearly Gondwanan and the Azores Laurasian. The Atlas mountains in northwest Africa are made up of several aggregated microdomains.
On the northeast, the major Arabian domain has separated off in the past and come back into contact, ending up with the triangular Sinai microdomain jammed in the gap. Arabia, which impacted with Laurasia to form the Zagros mountains, is shown by the plant distributions to be a Gondwanan domain. But it probably separated off relatively early, as it has its own genera within the common families, such as the carob, Ceratonia siliqua, in the legumes. It even has its own families, such as the pomegranate, Punicaceae, with only one genus and two species, one of which is confined to the tiny island of Socotra in the Gulf of Aden.
India was indisputably part of Gondwanaland, and positioned against the east African domains, but finding its exact former position is complicated by the fact that these domains have been shifted south, possibly to differing extents.
Madagascar was probably positioned against where Mozambique is now, but again with the complication that the northeast half of Mozambique may have been shifted south relative to the southwest half. Madagascar has coffee species, which fits in well with the pattern. Another excellent 'tie-in' genus is Adansonia, the boab or baobab family. This has one species growing right across north-tropical Africa and down the east coast, extending also to India (supposedly 'introduced by Arab traders'!), and a second species native to the Kimberly area of northwest Australia. But the big concentration, some 8-10 species, is in Madagascar.
Application of rubber-banding to Madagascan species will reveal its former connections, and because the island has many unique species related distantly or closely to those elsewhere, it should also be possible to work out when the different splits occurred, whether anciently or more recently. Of course the various edges of this domain were probably formed at different times.
The major link across from east Africa and the Mascarenes through India, southeast Asia, Indonesia and across the Pacific to Central America has already been shown. An excellent subject for rubber-banding is provided by the Seychelles islands north of Madagascar. These tiny, scattered islands include some with towering granite cliffs, virtually unique for isolated oceanic islands, and clear evidence of their 'continental' origin.
 Fig. 7.3. A Coco-de-Mer nut
Fig. 7.3. A Coco-de-Mer nut
The Seychelles has a number of endemic species (confined to that area), in the genera Acalypha, Begonia, Carissa, Dillenia (elephant apple), Diospyros (persimmon), Eugenia, Ficus (fig), Grewia, Pandanus (screwpine), and Pittosporum, all of which are typical of the major link. It also has the unique Double Coconut palm or Coco-de-Mer, Lodoicea, which has the largest seed in the world, weighing up to 20kg. It takes seven years for the fruit to mature, three years to germinate [49]. The nut bears an astounding resemblance to portions of the human female anatomy (Fig. 7.3), so much so that observers could quite understandably assume that some cosmic prankster has been at work!
North America
We have already identified some of the domains which make up the present North American continent, which is of medium complexity. The major domainographic features are the marked movements of shuffle belts or other domain aggregates north, on both sides of the continent.These shifts have carried palms up into California and Florida, and representatives of the Sapotaceae up into Florida (Fig. 6.1). Florida also has representatives of the Gondwanan custard-apple family, the Annonaceae. These include both Annona itself (the genus of the soursop, cherimoya, sugar apple, sweetsop, and most other custard apples) and also a genus restricted to North America, Asimina.
This contains several species, but the only one well known, even in North America, is the American Banana or Pawpaw (this is quite unrelated to the papaya, Carica papaya, which is also called a pawpaw in some parts of the world). The Asimina is said to be the largest native fruit in North America, good specimens weighing up to 500g. This again is a reflection of its origin, the larger fruits all originated in Gondwanaland -- we will go into the reason for this later.
Asiminas grow over wide areas of North America, principally on the Mississippi domain and the Atlantic domain (the coast from Florida to New York). Their distribution is evidence that the genus was split off from the rest of the family some time ago. The boundary between Asimina and Annona is reasonably distinct, running across northern Florida, and indicates that Florida itself is a fairly recently accreted island domain.
Other domains which could be distinguished in North America include the Rockies, western Mexico, Laurentia and three or four other large areas in continental Canada. Presence of the Great Lakes shows interaction of a mess of microdomains, but the most complex area of all is the west coast, which will take much study to sort out in detail.
Alaska is a complex of microdomains, as are the islands of northern Canada and, beyond them, Greenland. The Aleutian islands, stretching from Alaska right across the North Pacific to Asia, provide a very graphic illustration of fragments falling from the movement of the huge hinge formed as the North Pacific opened up. The Aleutians, and the Alaska Peninsula with which they terminate, appear to parallel Tierra del Fuego and the islands and submarine ridges which sweep down from South America to Antarctica, in that they represent the oldest ends of a microdomain shuffle belt. Both these shuffle belt ends show divergence from the usual equatorial-flight patterns, and indicate that this pattern of behaviour may not always apply in high latititudes.
Proposition 7F
Propositions relating to equatorial flight of domains may
lose validity at high latitudes
North America also shows a similar pattern to South America, in that plants native to the west coastal strip appear to 'intervene' between those east of the Rockies and the relatives of the latter across the Pacific. There is a strong link between the Texas-southern Mississippi flora and those of eastern Asia which appears to 'jump' over the west coast flora. This can be seen in the distribution maps of Pistacia (Fig. 4.10) and Carya (Fig. 4.11). The 'jump' is very obvious in Figure 7.4, from Berry [9], showing the distribution of the magnolia family.
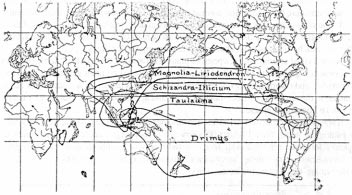 Fig. 7.4 Distributions of genera of the Magnolia family
Fig. 7.4 Distributions of genera of the Magnolia family
Try explaining the distributions in Figure 7.4 on the basis of migration across Bering Strait land bridges! There are also many other examples in Berry [9], often with distribution maps, showing the same pattern with such diverse genera as Tulip tree (Tupilifera), Sweetgum (Liquidambar), Tupelo (Nyssa), and Sassafras.
This pattern is not confined to plants. The existence of relational links between the fossil dinosaurs of China and western North America is well known, and is the subject of current study. What is interesting is that the American dinosaur occurrences are mostly east of the Rockies, especially in the Canadian province of Alberta.
Of course all these points confirm that the western shuffle belt has moved north along the coast, driving a wedge between the main North American flora and their counterparts at the same latitudes on the Asian coast.
Europe
Europe is a mess. The only major domain which it contains, covering the USSR west of the Urals and the North European Plain over as far as Germany, is structurally part of Asia (although it has undergone some re-seaming, along the Central Russian Uplands and the Volga Heights). Everything else is accreted and distorted domains and microdomains.Running all along its south it has the scattered parts of the Tethyan Girdle -- the band of island domains left from the original split of the holodomain into Gondwanaland and Laurasia. European domains in this band include Spain, Italy, Greece, the Mediterranean islands, and on to Turkey and the Central Asian republics of the USSR south of the Caucasus. All these domains have predominantly Laurasian rather than Gondwanan flora, and so should probably be classed with Laurasia. Also probably included with them is Ireland.
Ireland has, in its south-west corner, native occurrences of the Strawberry-Tree, Arbutus unedo. This also occurs right across the Mediterranean, and other species in the genus extend the range eastwards into the USSR (along the Tethyan Girdle) and, in the usual pattern, across the Atlantic (including the Azores) into Mexico and southwest USA (there is a concentration of seven species in Mexico).
The inference is that Ireland once lay in this band, and in fact, Ireland does fit quite snugly against northwest Portugal. Britain is a most complex mishmash of microdomains, and France, Germany, central Europe and the Balkans are also much re-worked, the most prominent feature being the Alps, formed when Italy collided with the northern domains.
To the north, Denmark, Norway, Sweden and Finland clearly consist of several domains pushed together. The Scandinavian mountains show marked dissection through the action of glaciers, which brings us to an observation concerning the Ice Ages.
When looked at from the viewpoint of plant distributions, the whole of northern Europe is notable for the poverty of its plant species. It has almost no plant genera confined to its boundaries, and comparatively few endemic species. This makes the application of rubber-banding to the area more difficult than for most, and also raises the question of why the area should be botanically sparse.
It appears that the answer to the last question lies in the Ice Ages. These were very recent in geological terms, only 10-110,000 years ago (0.01-0.11 my). There seems no doubt that glaciers formerly extended well south of their present European distribution, reaching southern Britain, for example. Inevitably, a consequence of being overrun by massive glaciers is the wiping-out of plant life in the area overrun.
Proposition 7G
Northern Europe has a relatively low level of plant diversity
because much of it was cleared of living plants through the action
of glaciers
Of course, when the glaciers retreated, vast areas of blank ecological niches were exposed, and plants came in to colonise these niches reasonably quickly. But because the ice ages were not so very long ago -- not only within the era of the species of man, but butting right up against the development of civilization -- there has just been insufficient time to develop the diversity of species and families which exist elsewhere.
It might be asked why the same situation does not exist across the whole of the temperate parts of the Northern Hemisphere. To some extent it does, in that colder areas are relatively poorer in plant diversity, but there is also another possibility. That is, that the recent ice-age glaciers were not centered on the North Pole (as is usually implicitly assumed), but were displaced more onto the European side for some reason.
Proposition 7H
Ice-age glaciers were not centered on the North Pole, but had
an area of influence displaced over into northern Europe
Asia
We have already seen that all the domains along the southern boundary of present-day Asia -- including Arabia, India, Indochina, and southern China -- are Gondwanan 'borrowings' since accreted to Laurasia. The Iran-Afghanistan domain is probably also one of these borrowings. The Tethyan Girdle which shows up in the Americas and Europe apparently either was never formed or was absorbed. There is a parallel formation, which includes the island domains and peninsulas of Indonesia, Malaya, and the Philippines, but in this the Gondwanan influence is predominant.In the main mass of Asia, north of the Gondwanan components, extensive re-seaming and shifting of domains has taken place, especially in south and west Siberia and in China. These actions have given rise to the many mountain ranges and to the strings of long lakes -- such as Baikal and Balkhash -- and to the Turfan Depression in northwest China.
 Fig. 7.5. Needles, cones and nuts of the
Siberian Stone Pine
Fig. 7.5. Needles, cones and nuts of the
Siberian Stone Pine
The Siberian area is notable for having the largest isocon envelopes currently existing on our planet. Stretching across the vast plains are huge areas in which the ecological conditions are relatively uniform, and so a particular plant species may come to have a very wide extent. One of the most prominent plants in this respect is, of course, a nut tree, the Siberian Stone Pine (Pinus sibirica).
This plant extends densely over more than 50 million hectares [71]. The potential for producing nuts is huge, as a large tree may produce 1500 cones each year. The average biological harvest of nuts from each hectare reaches 800 kg, so the potential annual yield of nuts is 40 million tonnes. This is many times the total weight of all nuts which figure in annual world trade.
From the domainographic viewpoint, the most interesting area in Asia is the Pacific Coast. This shows extensive evidence of domain flight. Japan and Korea, though rich in Laurasian flora such as oaks (Quercus), also have Gondwanan components such as yams (Dioscorea) and cycads, which have clearly come up from much further south. Both show the typical north-south oriented island or peninsula form, as do Sakhalin and Kamchatka to their north.
Southern China is one of the most floristically diverse regions of the world, with dense intermixtures of Laurasian and Gondwanan components. Any proposed boundary line would be open to argument, but it could lie somewhere along the line of the Nan Shan range.
As elsewhere, extensive equatorial flight has taken place along this coast. The whole of Manchuria and southeast USSR has apparently shifted north. The position is made more complex by the fact that domains of every size have been involved; we already saw in NU005 (Fig. 5.3) how the northeast USSR coast was a mass of microdomains. Instead of a straightforward shuffle belt like that of South America, we have a great mingling and jostling of large and small domains.
Plant distributions reflect the northward movement. A good example is the genus Actinidia, which contains the kiwifruit, Actinidia deliciosa. The kiwifruit, once called the chinese gooseberry, is one of 40 species in the genus, which is distributed not only in China (and across to Burma) but also north, right up into Siberia, where there are some very cold-hardy species. As well as Actinidia, the Actinidaceae family contains the genus Clematoclethra, with 10 species confined to China, and the big genus Saurauia.
In Asia Sauraia overlaps the southern range of Actinidia, extending through Burma (six species) as far east as Nepal. But Sauraia is also well represented in America, with species running from southern Mexico through Central America and right down the west coast as far as Chile. The genus is absent from the Caribbean and almost absent from the Guianas and Brazil. Again this is an absolutely typical pattern, the family showing strong links across the north equatorial Pacific, and strong equatorial flight, northwards in Asia and southwards in America.
The Polar Regions
Because of the paucity of living plants in the polar regions, it is hard to apply standard rubber-banding techniques to these areas. However, some information can be got from studies of fossil plants and of animal distributions. Although difficult to apply, studies of marine flora and fauna may also be productive.Having looked at where domain movements have taken place around the planet, we are now in a position to move on to the effects of these movements on the local landscapes -- how they got their ups and downs.

(Full list of references at NURefs)
[9]. Edward W Berry. Tree ancestors. Williams & Wilkins, Baltimore, 1923.
[38]. Alex Harris. Spiderman links blind hunter to lost continent. The West Australian/ Oct 4, 1988.
[39]. Dennis A Hearne. Some nut trees of the Northern Territory. West Australian Nutgrowing Society Yearbook/ 2 p43, 1976.
[49]. Guy Lionnet. The double coconut of the Seychelles. West Australian Nutgrowing Society Yearbook/ 2 p6-20, 1976.
[52]. John D Mitchell. The cashew and its relatives. New York Botanical Garden: Memoir 42, 1987.
[71]. A T Savel'ev. Nedrevesnaya produktsiya lesa [Non-timber forest products]. Lesprom, Moskva, 1980.
NU008: Making Mountains Out Of Movements
NU006: More Distributions And Evidence
Go to the NUSite Home Page
Version 2.0, 2004, PDFs etc on World Wide Web (http://www.aoi.com.au/matrix/Nuteeriat.htm)
Version 3.0, 2014 Sep 21, Reworked from Chapter 7 of "Nuteeriat" as one article in a suite on the World Wide Web.